В. В. Ашапкин, к. б. н., с. н. с., НИИ физико-химической биологии им. А. Н. Белозерского, Московский государственный университет им. М. В. Ломоносова
Е. Н. Волкова, д. м. н., профессор, директор Научно-образовательного департамента компании Premierpharm
Л. И. Кутуева, м. н. с., НИИ физико-химической биологии им. А. Н. Белозерского, Московский государственный университет им. М. В. Ломоносова
Б. Ф. Ванюшин, д. б. н., профессор, член-корр. РАН, заведующий отделом, НИИ физико-химической биологии им. А. Н. Белозерского, Московский государственный университет им. М. В. Ломоносова
Известно, что инъекционный препарат MesoSculpt C71™ на генном уровне способствует уменьшению массы белой жировой ткани [1]. Это уменьшение достигается за счет нескольких эффектов. Во-первых, препарат ингибирует экспрессию некоторых ключевых факторов адипогенеза (дифференцировки преадипоцитов в адипоциты), тем самым препятствуя росту жировой ткани за счет увеличения количества клеток (гиперплазии). Во-вторых, мезоскальп ингибирует синтез ряда ферментов липогенеза (накопления жиров в существующих адипоцитах), тем самым препятствуя росту жировой ткани за счет увеличения размеров индивидуальных клеток (гипертрофии). Наконец, он способствует активному уменьшению массы жировой ткани, стимулируя синтез ферментов, расщепляющих ранее накопленные липиды (триглицериды жирных кислот в липидных каплях). В данной работе с помощью исследования активности генов, продукты которых непосредственно участвуют в метаболизме липидов или регулируют различные его звенья, мы попытались оценить весомость вклада каждого из ранее описанных механизмов в общий баланс липидов, а также выяснить наиболее вероятные пути утилизации жирных кислот, освобождающихся из липидных капель при индуцированном препаратом гидролизе триглицеридов.
В качестве экспериментальной модели дифференцировки адипоцитов использовали иммортализованную клеточную линию мыши 3T3-L1, способную при соответствующих условиях дифференцироваться в адипоциты. Клетки культивировали в среде Игла модифицированной по Дальбеко, DMEM (Dulbecco’s Modified Eagle’s Megium, ПанЭко), содержащей 10 % телячьей эмбриональной сыворотки FBS (HyClone), 2мМ L-глутамина и 10мМ HEPES, pH 7,2–7,4, при температуре 37 ºC в увлажненной атмосфере 5 % CO2 в пластиковых чашках Петри. Для индукции дифференцировки к клеткам, достигшим 70–80 % конфлюентности добавляли изобутилметилксантин (IBMX) и дексаметазон до конечных концентраций 0,5мM и 1мкM, соответственно, на 48 часов. Для оценки влияния MesoSculpt C71™ на дифференцировку преадипоцитов его добавляли в культуральную среду до конечной концентрации 0,02 % на 1,5 часа, после чего клетки отмывали и продолжали выращивать в обычной среде. Смену среды делали каждые 2 дня. Для оценки влияния препарата на зрелые адипоциты его добавляли на 1,5 часа в конце процесса дифференцировки (на 8 сутки). Затем клетки отмывали и культивировали еще 24 часа. Контрольные клетки дифференцировали аналогичным образом, но без добавления MesoSculpt C71™.
По окончании дифференцировки среду удаляли, а к клеткам добавляли реагент, стабилизирующий РНК, RNAprotect Cell Reagent (Qiagen, Германия). После открепления клеток от поверхности чашек Петри под действием реагента образующуюся суспензию переносили в стерильные пластиковые микропробирки и хранили несколько суток в холодильнике до выделения РНК.
Выделение и очистку суммарной РНК из клеток осуществляли с помощью набора RNeasy Mini Kit (Qiagen, Германия) по прописи, рекомендованной фирмой-производителем набора. Полученные образцы РНК использовали для синтеза первой цепи кДНК с помощью набора обратной транскрипции Maxima First Strand cDNA Synthesis Kit (Thermo Scientific, США) по прописи, рекомендованной фирмой-производителем набора. В качестве матрицы на каждую реакцию обратной транскрипции объемом 20 мкл использовали 200 нг очищенной суммарной РНК. Полученную реакционную смесь использовали непосредственно как матрицу для ПЦР из расчета 1 мкл смеси на реакцию объемом 25 мкл. Количественную ПЦР с флуоресцентно мечеными гибридизационными зондами (TaqMan qPCR) проводили с помощью набора qPCRmix-HS (Евроген, Россия) и термоциклера ДТ-322 (ДНК Технология, Россия). В качестве внутреннего стандарта использовали кДНК референсного гена Gapdh. Ее концентрацию измеряли в тех же самых ПЦР смесях, используя меченый другой флуоресцентной меткой гибридизационный зонд: динамику амплификации кДНК для исследуемых генов измеряли по росту флуоресценции в канале Fam, а динамику амплификации для референсного гена – в канале Hex. Полученные данные импортировали в программу Microsoft Excel 2003 и обрабатывали статистически, принимая концентрацию мРНК референсного гена во всех образцах за 1. Для каждого образца проводили минимум три параллельные ПЦР в соседних лунках прибора. Конструирование олигонуклеотидных праймеров и гибридизационных зондов для количественной ПЦР осуществляли с помощью онлайн-сервиса IDT PrimerQuest (https://eu.idtdna.com/PrimerQuest/Home/Index), а их синтез – в компании Синтол (Россия).
Исследование динамики дифференцировки по соотношению размеров клеток и их гранулярности [2] и с помощью липофильного красителя Nile Red осуществляли на проточном цитофлуориметре Beckman-Coulter FC500, используя аргоновый лазер (488 нм). Ультраструктуру залитых в Эпон клеток исследовали на ультратонких срезах, окрашенных уранил-ацетатом и цитратом свинца под электронным микроскопом JEM-1400 (JEOL, Япония).
На рисунке 1 представлена схема адипогенеза в белой жировой ткани млекопитающих, на которой указаны ключевые регуляторные факторы, исследованные в данной работе.
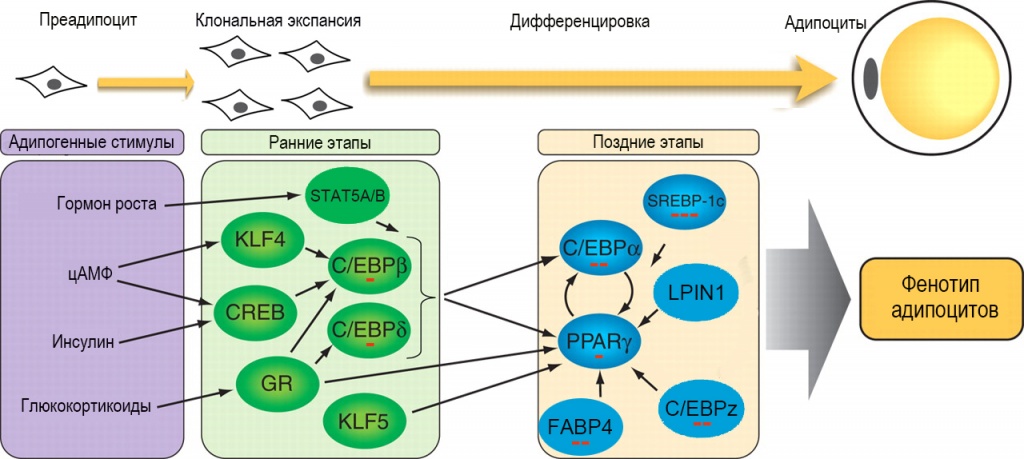
Рис. 1. Факторы регуляции адипогенеза. Зеленым цветом выделены факторы раннего адипогенеза, активируемые адипогенными гормонами и цАМФ. В свою очередь они индуцируют синтез факторов позднего адипогенеза (выделены синим цветом), контролирующих окончательную дифференцировку и поддержание стабильного фенотипа зрелых адипоцитов. KLF – Krüppel-like factor, CREB – cAMP response element binding protein, C/EBP– CCAAT/enhancer binding protein, GR – glucocorticoid receptor, STAT5A/B – signal transducer and activator of transcription 5A/B, PPARγ – peroxisome proliferator-activated receptor γ, FABP4 – fatty acid binding protein 4, LPIN1 – lipin 1, SREBP-1c - sterol regulatory element binding protein 1c. Красными минусами указаны обнаруженные эффекты мезоскальпа на экспрессию соответствующих генов: один минус – слабое ингибирование (в <2 раза), два минуса – умеренное ингибирование (2-5 раз), три минуса – сильное ингибирование (>5 раз)
Побудительными стимулами, запускающими процесс дифференцировки, являются гормональные факторы (глюкокортикоиды, инсулин и гормон роста) и агенты, вызывающие увеличение концентрации цАМФ [3]. Полный цикл адипогенеза занимает 10–12 дней и состоит из двух этапов, каждый из которых контролируется своей особой группой регуляторных факторов. На раннем этапе происходит деление (клональная экспансия) преадипоцитов. Главную роль в этом процессе играют факторы транскрипции C/EBPβ и C/EBPδ. Синтез каждого из них ингибируется мезоскальпом в 1,5–2 раза. По окончании клональной экспансии происходит фосфорилирование молекул C/EBPβ и C/EBPδ, в результате которого они приобретают способность индуцировать синтез факторов позднего адипогенеза. Решающую роль в позднем этапе адипогенеза и поддержании стабильного фенотипа дифференцированных адипоцитов играют мастер-регуляторы адипогенеза, транскрипционные факторы PPARγ и C/EBPα, объединенные в единую регуляторную систему взаимными положительными обратными связями. Наиболее важными «помощниками» мастер-регуляторов являются транскрипционные факторы C/EBPz и SREBP-1c, белок транспорта жирных кислот FABP4 и коактиваторный белок липин 1 (LPIN1, также известный как фосфогидролаза фосфатидных кислот). Мезоскальп оказывает слабовыраженное ингибирующее действие на активность гена PPARγ, умеренно ингибирует экспрессию генов CEBPα, CEBPz и FABP4 и сильно – гена SREBP-1c. Очевидно, MesoSculpt C71™ в значительной степени подавляет адипогенез. Следует подчеркнуть, что транскрипционный фактор SREBP-1c, помимо участия в контроле адипогенеза, играет роль в жизни зрелых адипоцитов как мастер-регулятор экспрессии генов липогенеза. Вполне вероятно, что обнаруженное сильное ингибирование экспрессии гена SREBP-1c мезоскальпом является первичным эффектом этого препарата, а уменьшение экспрессии генов липогенеза, которое будет обсуждаться ниже, вторичными эффектами. Цитофлуориметрическое исследование подтвердило вывод об ингибировании адипогенеза мезоскальпом: добавление препарата одновременно с индукторами дифференцировки почти полностью блокировало образование адипоцитов, а его добавление на поздних этапах дифференцировки значительно (на 35 %) уменьшало количество липидов, накопившихся в клетках к концу дифференцировки. Электронно-микроскопическое исследование показало, что при добавлении мезоскальпа на ранних этапах дифференцировки клетки сохраняют морфологические признаки, характерные для преадипоцитов (незначительное содержание липидных капель (ЛК), много митохондрий) даже к концу периода дифференцировки (рис. 2, А и Б). При добавлении препарата на завершающих этапах дифференцировки образуются клетки, более морфологически сходные с нормальными зрелыми адипоцитами, но содержание ЛК в них относительно невелико (рис. 2, В и Г).
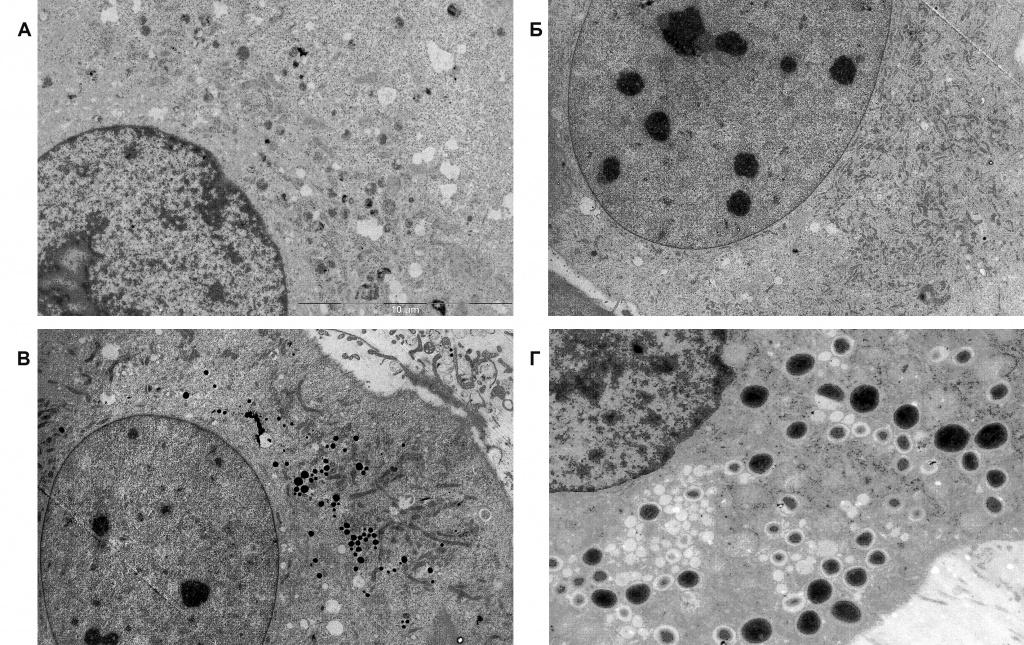
Рис. 2. Ультраструктура клеток, индуцированных к дифференцировке в разных условиях. А – фрагмент преадипоцита перед добавлением индукторов дифференцировки; немногочисленные липидные капли (ЛК) в цитоплазме (белые пятна); Б – адипоцит после дифференцировки с добавлением мезоскальпа на ранних этапах; немногочисленные ЛК среди множества мелких митохондрий; В – адипоцит после дифференцировки с добавлением мезоскальпа на поздних этапах; содержание ЛК (белые и черные пятна округлой формы) несколько выше, чем в преадипоцитах (А), но существенно ниже, чем в контрольных адипоцитах (Г); Г – адипоцит после дифференцировки в стандартных условиях (без добавления мезоскальпа); многочисленные и более крупные ЛК (белые и черные пятна округлой формы), редкие митохондрии
В зрелых адипоцитах прогрессивное накопление липидов (липогенез) в первую очередь происходит в результате захвата жирных кислот (ЖК) из кровяного русла. Наиболее удобной формой хранения и транспортировки жирных кислот являются триглицериды (ТГ). Однако сами ТГ не способны проникать через клеточную мембрану, поэтому для переноса внутрь клетки они должны быть гидролизованы специальным ферментом, липопротеинлипазой (lipoprotein lipase, LPL) [4–6]. В просвете капилляров богатые ТГ липопротеины пищевых хиломикронов или образующихся в печени липопротеидов очень низкой плотности (VLDL) расщепляются липопротеинлипазой, а образующиеся при этом свободные ЖК транспортируются внутрь адипоцитов с помощью специального белка, транслоказы ЖК (FAT – Fatty acids translocase, известного также как CD36) (рис. 3).
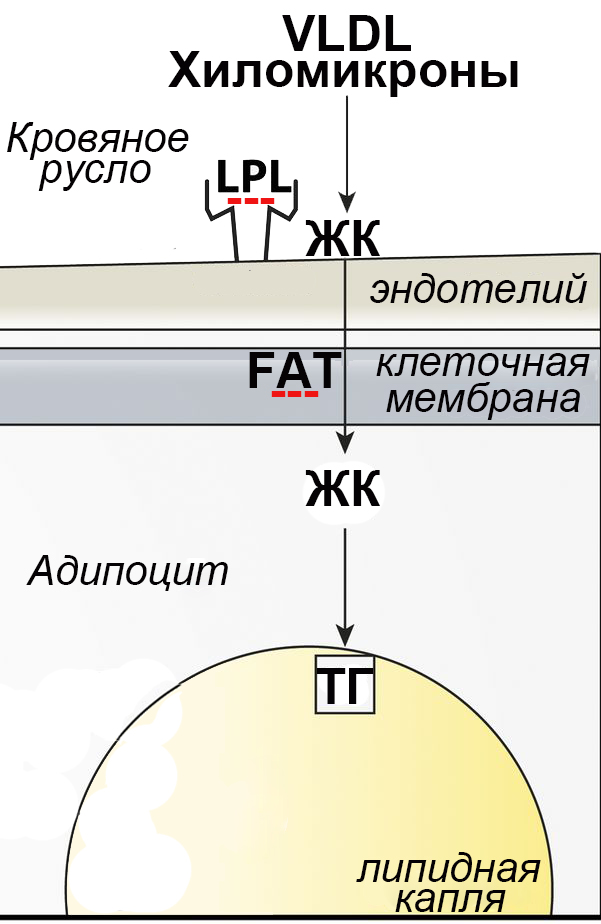
Рис. 3. Гидролиз ТГ на внутренней поверхности капилляров и транспорт ЖК в адипоциты. Молекулы липопротеинлипазы (LPL), синтезируемые в адипоцитах, транспортируются в просвет капилляров и удерживаются на их внутренних стенках с помощью специального гликопротеина. Освобождающиеся при гидролизе ТГ хиломикронов и липидов очень низкой плотности свободные ЖК транспортируются через мембрану адипоцитов белком FAT. В цитоплазме происходит их повторная этерификация и включение образующихся ТГ в липидные капли. Тремя красными минусами показаны обнаруженные сильные (в > 5 раз) ингибирующие эффекты мезоскальпа на экспрессию генов LPL и FAT
Увеличение массы жировой ткани при потреблении богатой жирами пищи в значительной степени определяется именно активностью LPL и FAT. В опытах на трансгенных мышах показано, что усиление экспрессии гена LPL приводит к увеличению массы белой жировой ткани, а ее ослабление – к уменьшению [4]. При инактивации гена FAT наблюдается резкое возрастание концентрации свободных ЖК в плазме крови и ингибирование активности LPL. Ранее мы уже описывали сильный ингибирующий эффект мезоскальпа на экспрессию гена LPL в адипоцитах [1]. В данной работе мы обнаружили, что примерно в такой же степени (в ~5 раз) препарат ингибирует и экспрессию гена FAT. Очевидно, что общим результатом этих двух ингибирующих эффектов должно быть сильно выраженное подавление захвата экзогенных жирных кислот адипоцитами. Одним из побочных эффектов такого подавления может быть резкое усиление захвата глюкозы адипоцитами [7]. Это в свою очередь могло бы стимулировать синтез жирных кислот de novo в самих адипоцитах. Поэтому мы исследовали экспрессию генов, кодирующих ключевые ферменты синтеза жирных кислот (рис. 4).
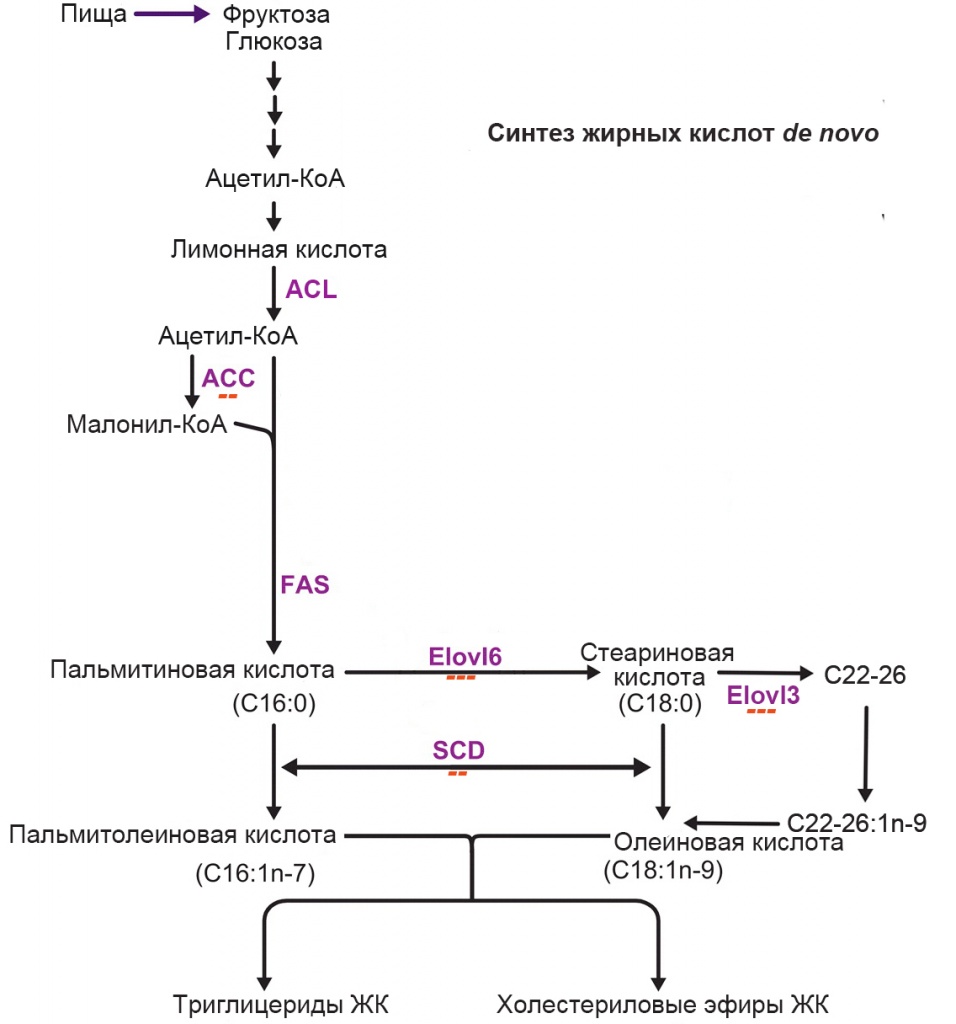
Рис. 4. Пути синтеза жирных кислот de novo. Пурпурным шрифтом обозначены ферменты, катализирующие последовательные этапы синтеза: ACL – ATP-citrate lyase, ACC – acetyl-CoA carboxylase, FAS - fatty acid synthase, Elovl – elongation of very long chain fatty acids, SCD – stearoyl-CoA desaturase. Красными минусами указаны обнаруженные эффекты мезоскальпа на экспрессию соответствующих генов: один минус – слабое ингибирование (в <2 раза), два минуса – умеренное ингибирование (2-5 раз), три минуса – сильное ингибирование (>5 раз)
Оказалось, что MesoSculpt C71™ сильно (в >5 раз) ингибирует экспрессию генов Elovl3 и Elovl6, умеренно – экспрессию генов ACC и SCD, и не влияет на экспрессию генов FAS и ACL. Следовательно, он в значительной степени подавляет синтез насыщенных и мононенасыщенных жирных кислот из внутренних источников.
Накопление больших количеств триглицеридов в адипоцитах белой жировой ткани сопровождается постепенным слиянием мелких ЛК во все более крупные (рис. 5).
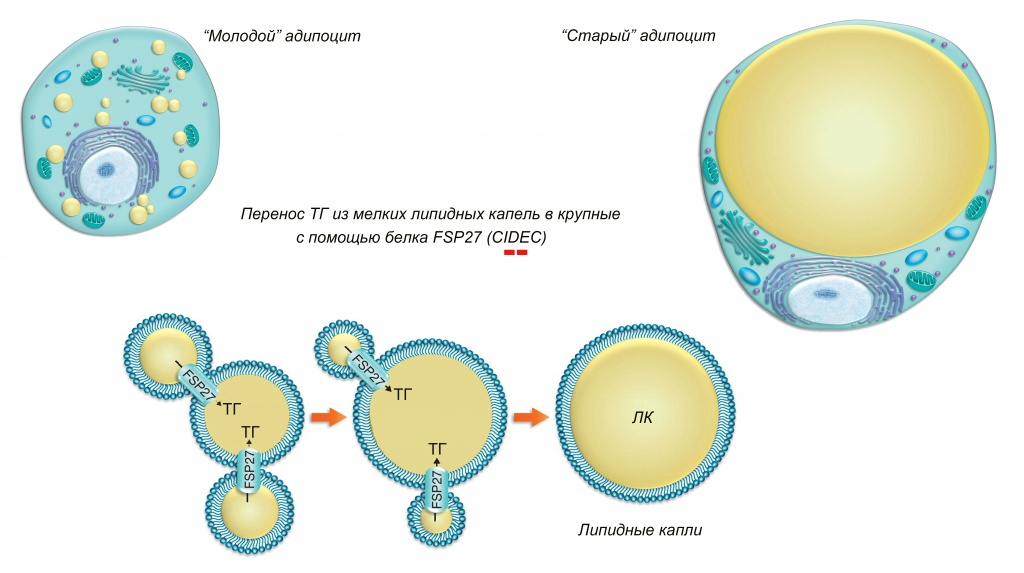
Рис. 5. Слияние липидных капель (ЛК) в результате «поглощения» мелких более крупными. FSP27 (CIDEC) – fat specific protein 27 (cell death-inducing DFF45-like effector C). Два красных минуса указывают на обнаруженное умеренное ингибирование экспрессии гена FSP27 (CIDEC) мезоскальпом
Важнейшую роль в этом процессе играют белки семейства CIDE [8]. Считается, что они образуют своеобразные ТГ-проводящие каналы в местах контакта разных ЛК, через которые происходит перенос липидов из одной в другую. При этом направление переноса зависит от соотношения различных белков данного семейства. В белых адипоцитах преобладающим является белок CIDEC, обеспечивающий транспорт липидов из мелких ЛК в более крупные вплоть до образования одной огромной ЛК (в унилокулярных адипоцитах белой жировой ткани диаметр ЛК может достигать 100 мкм и более). Такой вариант (максимальный объем при минимальной поверхности) оптимален для хранения липидов. В бурых адипоцитах преобладает белок CIDEA, обеспечивающий формирование мультилокулярной структуры, оптимальной для активной утилизации липидов. При индукции образования бежевых адипоцитов, например, в результате адаптации к холоду, активируется экспрессия CIDEA и унилокулярная морфология белых адипоцитов сменяется мультилокулярной. Результатом является многократное увеличение площади контакта между липидными каплями и цитоплазмой, что, естественно, способствует более активному липолизу и дальнейшей утилизации освобождающихся жирных кислот. Препарат MesoSculpt C71™ уменьшает уровень экспрессии гена CIDEC примерно в 4 раза, что явно должно препятствовать накоплению липидов в адипоцитах. К тому же, недавно обнаружено, что белок CIDEC ингибирует экспрессию гена ATGL [9] и активность самой липазы ATGL [10]. Следовательно, уменьшение экспрессии гена CIDEC должно приводит к дополнительному усилению липолиза за счет повышения активности ATGL.
Липолиз является процессом, противоположным липогенезу [11, 12]. При дефиците жирных кислот в активно утилизирующих их органах (миокард, скелетная мускулатура, печень), например, вследствие продолжительного голодания, происходит мобилизация запасенных в адипоцитах жирных кислот. При повышении тонуса симпатической нервной системы происходит стимуляции β-адренорецепторов на поверхности адипоцитов, активация синтеза цАМФ и зависящей от него протеинкиназы А (PKA). Эта протеинкиназа фосфорилирует гормон-чувствительную липазу HSL и белки, ассоциированные с ЛК, перилипины (рис. 6).
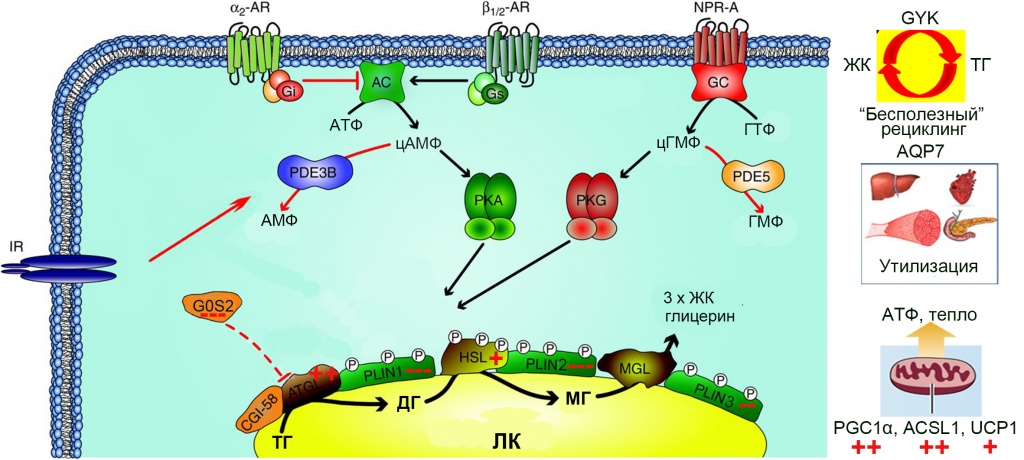
Рис. 6. Регуляция липолиза в адипоцитах белой жировой ткани. Черными линиями обозначены стимулирующие липолиз влияния, красными – ингибирующие. Справа показаны основные пути утилизации ЖК. AR – адренорецепторы; NPR-A – рецептор натрийуретических пептидов тип А; IR – рецептор инсулина; Gs – G-белок стимулирующего типа; Gi – G-белок ингибирующего типа; AC – аденилатциклаза; GC – гуанилатциклаза; PDE – фосфодиэстераза; PKA – протеинкиназа A; PKG – протеинкиназа G; ТГ – триглицериды; ДГ – диглицериды; МГ – моноглицериды; ATGL – ТГ липаза адипоцитов (adipose TG lipase); HSL – гормон-чувствительная липаза (hormone-sensitive lipase); MGL – МГ липаза (monoglyceride lipase); PLIN – перилипин; CGI-58 – comparative gene identification-58; G0S2 – G0/G1 switch 2; GYK – глицеролкиназа; AQP7 – аквапорин 7; PGC1α – peroxisome proliferator-activated receptor γ coactivator 1α; ACSL1 – ацил-КоА-синтетаза 1 (acyl-CoA synthetase-1); UCP1 – разобщающий белок 1 (uncoupling protein 1). Тремя красными минусами показано сильное ингибирование мезоскальпом экспрессии генов G0S2, PLIN1 и PLIN2, двумя – умеренное ингибирование гена PLIN3, красными плюсами – стимулирующие эффекты мезоскальпа на экспрессию соответствующих генов: один плюс – слабая стимуляция (в <2 раза), два – умеренная (2–5 раз)
Мощными активаторами липолиза являются натрийуретические пептиды. Они действуют через рецептор натрийуретических пептидов А, обладающий гуанилат-циклазной активностью. В результате происходит цГМФ-зависимая активация протеинкиназы G (PKG) и активирующее фосфорилирование липазы HSL и перилипинов. Концентрация натрийуретических пептидов в крови повышается в результате физических нагрузок. Повышение концентрации инсулина, например, после приема пищи, наоборот ингибирует липолиз. Инсулин активирует фосфодиэстеразу 3B, гидролизующую цАМФ и тем самым подавляет PKA-зависимое фосфорилирование. Активность протеинкиназы G подавляется в результате гидролиза цГМФ фосфодиэстеразой 5, но факторы, активирующие эту фосфодиэстеразу, пока не идентифицированы. Расщепление каждой молекулы триглицеридов происходит на поверхности липидной капли последовательно тремя липазами, ATGL, HSL и MGL. В результате образуется три молекулы ЖК и одна молекула глицерина. Краткосрочная регуляция активности этих липаз осуществляется путем фосфорилирования HSL и перилипинов, в результате которого изменяется сродство ATGL к регуляторным белкам G0S2 и CGI-58, а также доступность поверхности ЛК для липаз и их каталитическая активность. Ключевую роль в этом процессе играет перилипин 1. В отсутствии стимуляции нефосфорилированные молекулы перилипина 1 прочно ассоциированы с белком CGI-58, являющимся мощным коактиватором ATGL. Сама ATGL при этом прочно ассоциирована с корепрессорным белком G0S2 и каталитически неактивна. Фосфорилирование перилипина 1 нарушает его ассоциацию с коактиватором CGI-58, в результате чего последний освобождается и, вытесняя корепрессор G0S2, активирует липазу ATGL. При этом происходит и активация всего липолиза в целом, поскольку именно отщепление первой молекулы ЖК от молекулы ТГ является лимитирующей стадией процесса [10]. Длительная адаптация белой жировой ткани к изменившимся условиям внешней или внутренней среды (недостаток пищи, продолжительное охлаждение, избыточные физические нагрузки) достигается путем устойчивых изменений в активности генов. MesoSculpt C71TM умеренно (в ~2 раза) стимулирует экспрессию гена ATGL, несколько слабее (в 1,7 раза) – экспрессию гена HSL, и не влияет на экспрессию гена MGL. Таким образом, наблюдается стимуляция липолиза ТГ. Еще более выраженной эта стимуляция становится благодаря сильному (>5 раз) ингибированию экспрессии генов перилипина 1 и корепрессорного белка G0S2. В опытах на трансгенных мышах показано, что выключение (knockout) гена PLIN1 приводит к сильно выраженному уменьшению массы белой жировой ткани в результате повышенного базового уровня липолиза [12]. Аналогично, выключение гена G0S2 приводит к выраженной стимуляции [13], а его избыточная экспрессия – подавлению липолиза [14]. Таким образом, умеренная стимуляция мезоскальпом экспрессии генов ATGL и HSL, наряду с подавлением экспрессии генов двух главных ингибиторов ATGL, должна весьма значительно стимулировать липолиз ТГ в адипоцитах белой жировой ткани. Мезоскальп также вызывает существенное (в >5 раз) подавление экспрессии гена PLIN2 и умеренное (в ~2 раза) – гена PLIN3. Показано, что в культуре клеток избыточная экспрессия гена PLIN2 приводит к подавлению липолиза ТГ на поверхности ЛК и уменьшает доступность поверхности ЛК для ATGL [15]. Инактивация гена PLIN2 у мышей делает их резистентными к ожирению, вызванному богатой жирами диетой, что коррелирует с увеличением доли бежевых адипоцитов в подкожной жировой ткани [16]. Перилипины 2 и 3 могут взаимно компенсировать недостачу друг друга на поверхности ЛК, а уменьшение их количества способствует доступу ATGL к поверхности ЛК и активации липолиза [17]. Механизмы этих эффектов пока мало изучены, но какими бы они ни были, ингибирование экспрессии генов PLIN2 и PLIN3 мезоскальпом должно дополнительно стимулировать липолиз ТГ. Перилипин 4 в больших количествах присутствует в белой жировой ткани и в меньших – в скелетных мышцах и сердце. В бурой жировой ткани он практически отсутствует [18]. Для перилипина 5 характерна противоположная локализация: его много в клетках с высокой окислительной активностью (бурая жировая ткань, сердце, скелетные мышцы, печень). При липолитической стимуляции адипоцитов агонистом β-адренорецепторов изопротеренолом наблюдается фрагментация ЛК на множество микрокапель, с поверхностью которых ассоциирован перилипин 4 [19]. И хотя функциональное значение этой ассоциации остается загадочным, фрагментация ЛК как таковая очевидно способствует активации липолиза ТГ, увеличивая отношение площади поверхности ЛК, доступной для липаз, к их объему. Мезоскальп заметно (в 4–5 раз) увеличивает уровень экспрессии гена PLIN4, что, вероятно, приводит к стимуляции липолиза. В отличие от перилипина 4 и аналогично перилипину 1 в белой жировой ткани, перилипин 5 в экспрессирующих его клетках препятствует липолизу ТГ в базовом нефосфорилированном состоянии, но этот барьер исчезает при его фосфорилировании протеинкиназой A [20, 21]. В молекуле перилипина 5 обнаружен участок связывания с митохондриями, обеспечивающий прямой физический контакт между этими органеллами и ЛК [22]. Вполне вероятно, что именно эта особенность обуславливает избирательную экспрессию перилипина 5 в бурых адипоцитах и других клетках с высокой оксидативной активностью. Мезоскальп незначительно (в ~ 1,3 раза) повышает уровень экспрессии гена PLIN5. Возможно, это отражает индуцированную препаратом трансдифференцировку части белых адипоцитов в бежевые и объясняет наблюдавшуюся нами с помощью ЭМ тесную ассоциацию ЛК с митохондриями при действии препарата (рис. 7).
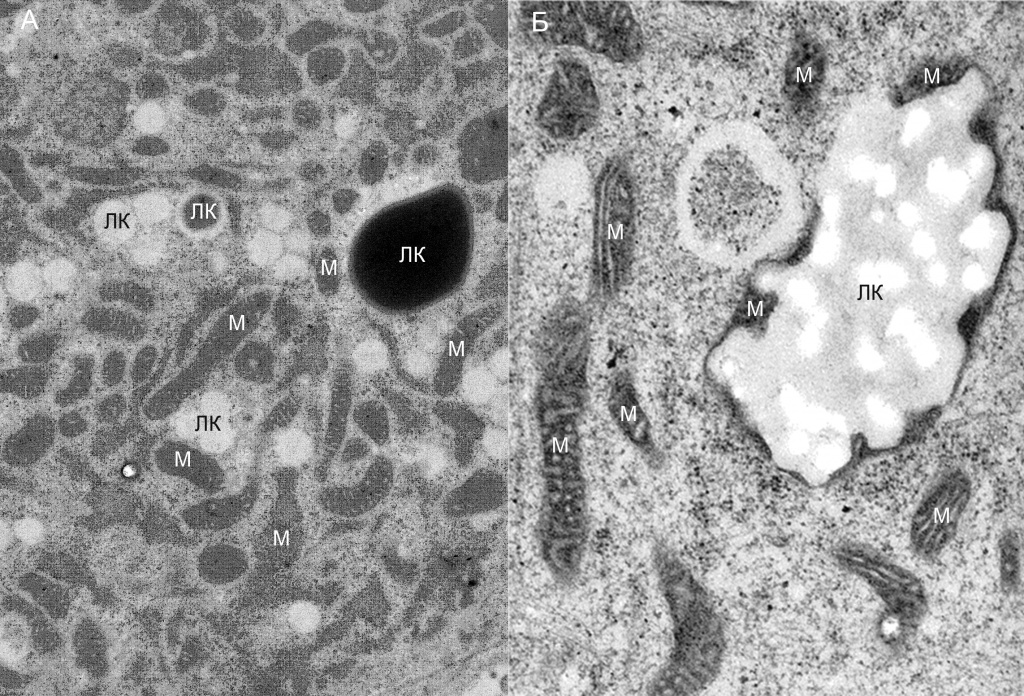
Рис. 7. Тесная физическая ассоциация ЛК с митохондриями при действии мезоскальпа. А – ЛК окружены множеством митохондрий (M). Различные размеры и окраска ЛК и митохондрий обусловлены их варьирующей ориентацией в трехмерном пространстве клеток по отношению к плоскости среза; Б – ЛК, окруженная митохондриями, при большем увеличении
Одним из главных молекулярных маркеров активного образования новых митохондрий является белок PGC1α [23]. Мы наблюдали умеренную (в ~2 раза) стимуляцию экспрессии гена PGC1α мезоскальпом, что явно свидетельствует об активации биогенеза (образования) митохондрий. Функциональным значением такой активации, по-видимому, является β-окисление ЖК, освобождающихся при индуцированном липолизе ТГ. Такое предположение естественным образом объясняет наблюдающуюся тесную ассоциацию между ЛК и митохондриями в обработанных препаратом адипоцитах. Оно подтверждается также наблюдавшейся нами стимуляцией (в ~2 раза) экспрессии гена ACSL1, которая, как известно, коррелирует с интенсивностью процесса β-окисление ЖК [24]. Таким образом, окисление ЖК в митохондриях является, как минимум, одним из способов утилизации ЖК, освобождающихся из ЛК в результате активации мезоскальпом процессов липолиза. Известно, что в митохондриях большинства клеток, в том числе адипоцитов белой жировой ткани, окисление ЖК сопряжено с синтезом АТФ, в то время как в адипоцитах бурой жировой ткани и подобных им бежевых адипоцитах окисление ЖК происходит без сопутствующего синтеза АТФ, а освобождающаяся при этом энергия рассеивается в виде тепла [25]. Белок UCP1, ответственный за разобщение процессов окисления и синтеза АТФ, является главным молекулярным маркером бурых и бежевых адипоцитов. Мы обнаружили, что ген UCP1 очень слабо экспрессируется в контрольных адипоцитах, а при добавлении мезоскальпа уровень его экспрессии возрастает в 1,3–1,8 раза, то есть в меньшей по сравнению с генами PGC1α и ACSL1 степени. Логично предположить, что наблюдаемая активация β-окисления ЖК происходит как за счет увеличения количества митохондрий в самих белых адипоцитах, так, отчасти, и за счет трансдифференцировки какой-то части белых адипоцитов в бежевые. Наиболее эффективно, с точки зрения «сжигания» избыточных жиров, окисление ЖК в бежевых адипоцитах, поскольку при этом высвобождаемая энергия не запасается в виде молекул АТФ (которые, в принципе, могут быть использованы и для синтеза новых молекул ТГ), а бесследно рассеивается в форме тепла. Тем не менее, и обычная утилизация ЖК в собственных митохондриях белых адипоцитов может быть вполне эффективным способом уменьшения массы белой жировой ткани [26]. Наконец, нельзя не упомянуть еще о двух возможных способах утилизации освобождающихся при липолизе молекул ЖК. Первый связан с повторной этерификацией ЖК и возвращением образующихся при этом ТГ в ЛК. На первый взгляд результат такого процесса все возвращает в исходное состояние, поэтому в англоязычной литературе он обозначается как «бесполезный» (futile) цикл. В действительности реэтерификация ЖК требует затрат довольно значительных количеств АТФ и в этом смысле вовсе не является бесполезной. Лимитирующим фактором в «бесполезном» цикле является активность фермента глицеролкиназы (GYK). Оказалось, что мезоскальп не изменяет экспрессии гена GYK, поэтому данный вариант утилизации ЖК, скорее всего, не играет существенной роли в механизмах его липолитического действия. Еще один возможный вариант утилизации ЖК – их освобождение из адипоцитов в кровоток и доставка в другие органы. Оба процесса в значительной степени зависят от экспрессии гена AQP7, кодирующего мембранный белок водно-глицеринового канала. При низкой экспрессии этого гена образующиеся при липолизе молекулы глицерина в основном остаются внутри адипоцита, что приводит к активации утилизации ЖК и глицерина по пути «бесполезного» цикла [27]. Мы не наблюдали сколько-нибудь существенных изменений в уровне экспрессии гена AQP7 под действием мезоскальпа. Следовательно, эффективность утилизации ЖК за счет реэтерификации и экспорта остается на базовом уровне. Это в свою очередь означает, что основным способом утилизации ЖК при действии мезоскальпа является их β-окисление в митохондриях белых и бежевых адипоцитов.
Результаты описанного исследования подтверждают позиционирование инъекционного препарата MesoSculpt C71™ как средства, уменьшающего массу подкожных жировых отложений. При этом механизмы действия препарата множественные. Одним из наиболее выраженных является ингибирование захвата ЖК из пищевых источников за счет подавления активности генов липопротеинлипазы (LPL) и транслоказы ЖК (FAT). Этот механизм, очевидно, наиболее важен при богатой жирами диете. В случае избыточного потребления углеводов на первый план выходит ингибирование активности генов, кодирующих ферменты синтеза ЖК de novo. В обоих случаях увеличению жировой массы препятствует также ингибирование процессов адипогенеза. Наиболее выраженным эффектом, ответственным за уменьшение массы ранее накопленных жиров, является стимуляция процессов липолиза ТГ в адипоцитах белой жировой ткани и последующего β-окисления освобождающихся ЖК в митохондриях, тесно ассоциированных с ЛК. Стимуляция липолиза и последующей утилизации ЖК в свою очередь также достигаются за счет множественных эффектов. Во-первых, это – стимуляция собственно расщепления ТГ в результате увеличения экспрессии генов липазы ТГ (ATGL) и гормон-чувствительной липазы ДГ (HSL), а также их «помощника» перилипина 4. Во-вторых, наблюдается ингибирование генов, кодирующих белки, препятствующие липолизу (G0S2 и перилипины 1-3). Наконец, мы обнаружили активизацию β-окисления освобождающихся в результате липолиза ЖК в митохондриях. При этом наблюдается увеличение числа митохондрий в самих адипоцитах белой жировой ткани и, в какой-то степени, трансдифференцировка белых адипоцитов в бежевые, способные наиболее эффективно «сжигать» ЖК без сопутствующего образования АТФ. Судя по соотношению активностей генов, кодирующих маркеры биогенеза митохондрий (PGC1α), β-окисления ЖК в митохондриях (ACSL1) и образования бежевых адипоцитов (UCP1, PLIN5), β-окисление ЖК в собственных митохондриях белых адипоцитов является преобладающим путем их утилизации, а их «сжигание» в бежевых адипоцитах – вспомогательным. Следует оговориться, однако, что в наших экспериментах воздействие препарата на клетки было однократным и непродолжительным. Вполне вероятно, что при более длительном или повторном действии препарата MesoSculpt C71™ вклад альтернативного пути утилизации ЖК, связанного с образованием бежевых адипоцитов, будет более весомым. По своей природе, процесс трансдифференцировки белых адипоцитов в бежевые является эпигенетическим репрограммированием. Это означает, что возникшие бежевые адипоциты останутся таковыми и после окончания действия препарата. При его повторном действии количество бежевых адипоцитов будет увеличиваться, а следовательно, будет увеличиваться и их удельный вес в общем балансе утилизации ЖК.














 16766
16766


Комментарии